小泡的定向运输、停靠和融合机理[细胞生物学]
发布时间:2011-10-18 共1页
小泡的定向运输、停靠和融合机理
无论是选择性还是非选择性的运输小泡, 它们都必须高度选择性地有方向地到达目的地,那么定向运输和停泊的标志是什么呢? 到达目的地后如何停泊?各种小泡都是膜封闭的结构,它们又是怎样突破膜结构的障碍释放出内含物?
■ 运输小泡寻靶: SNARE 假说
James Rothman和他的同事根据对动物细胞融合研究的发现, 提出有关小泡寻靶的SNARE假说(SNARE hypothesis)。
● NSF和SNAPs
他们发现动物细胞融合需要一种可溶性的细胞质蛋白,叫做N-乙基马来酰亚胺敏感的融合蛋白(N-ethylmaleimide-sensitive fusion protein,NSF)以及其它几种可溶性的NSF附着蛋白(soluble NSF attachment protein,SNAPs)。NSF是一种四聚体,四个亚基都相同。SNAPs 有α-、β-和γ- SNAPs等。
● SNARE 假说
由于NSF/ SNAPs能够介导不同类型小泡的融合,说明它没有特异性。据此Rothman提出一种假说:膜融合的特异性是由另外的膜蛋白提供的,把这种蛋白称为SNAP受体蛋白(SNAP receptors),或称为SNAREs,这种蛋白可以作为膜融合时SNAPs的附着点。
● 不同的小泡具有不同的SNAREs
按照Rothman的SNARE假说,每一种运输小泡都有一个特殊的V-SNARE(vesicle-SNAP receptor)标志,能够同适当的靶膜上的T-SNARE(target-SNAP receptor)标志相互作用。一种运输小泡在没有找到合适的靶位点之前有可能同几种不同的膜位点进行过暂时性地接触,这种接触是不稳定的,只有找到真正的靶位点才会形成稳定的结构(图9-70)。
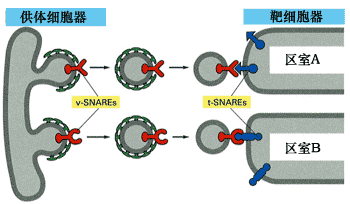
图9-70 运输小泡寻靶
不同的小泡上具有不同的V-SNARE, 它能识别不同靶膜上的T-SNARE并与之结合,以此保证运输小泡到达正确的目的地。
■ Rab蛋白(Rab protein)在小泡运输与融合中的调节作用[]
Rab蛋白家族是真核细胞中控制小泡转运的GTP结合蛋白。
● Rab蛋白是一类调节型的单体GTPase, 所有的Rab蛋白都是由大约200个氨基酸组成的,并且有类似于Ras蛋白的重叠结构。它能够结合GTP并将GTP水解,因此认为Rab蛋白通过GTP的循环来调节小泡的融合(图9-71)。
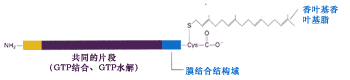
图9-71 Rab蛋白的结构
● Rab蛋白在小泡的转运和融合中的调节机理可能是:供体膜上的鸟嘌呤核苷释放蛋白(GNRP)识别胞质溶胶中特异的Rab蛋白,诱导GDP的释放并和GTP结合,进而改变Rab蛋白的构型,改变了构型的Rab蛋白暴露出其脂基团,从而将Rab蛋白锚定到膜上。运输小泡形成后,在V-SNARE的引导下,到达受体膜的T-SNARE部位,Rab帮助小泡与受体膜结合。Rab蛋白上的GTP水解后从膜中释放出来,而小泡却锁定在受体膜上,释放出的Rab进入胞质溶胶进行再利用(图9-72)。
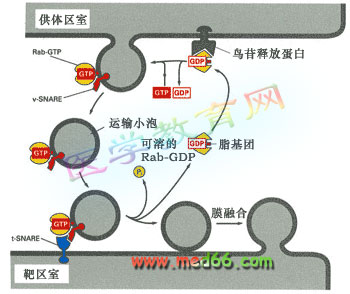
图9-72 Rab蛋白在小泡运输和融合中的调节作用
● Rab蛋白作用的实验证据
有一些离体实验支持Rab蛋白在小泡运输和融合中的作用。如Rab5定位于初级内体的膜上, 无细胞系统实验表明, 初级内体间的相互融合需要Rab5的存在, 也不能用其它类Rab蛋白取代。加入Rab和GTP后初级内体间就能发生融合, 说明Rab和GTP是初级内体融合的触发剂。同样发现Rab1蛋白是ER同高尔基体小泡所必需的。
■ 小泡融合模型
根据SNARE假说以及对Rab蛋白等生化和遗传学研究结果表明, 小泡融合时,需要V-SNARE、T-SNARE和融合蛋白SNAP25的存在。将含有纯化的V-SNARE的人工脂质体与含有T-SNARE/SNAP25复合物的脂质体一起温育,渐渐地两种膜融合到一起, V-SNARE、T-SNARE/SNAP25出现在同一种膜上。在细胞中融合只要几秒钟,但需要几种胞质溶胶蛋白的参与,包括NSF、α-、β-和γ-SNAPs,它们的作用可能是使V-SNARE、T-SNARE/SNAP25解离以便于再利用, 同时扩大融合。图9-73是根据体外实验的结果提出的小泡融合模型。

图9-73 运输小泡寻靶与融合模型
① 运输小泡通过小泡膜中的V-SNARE与靶膜T-SNRE/SNAP25复合物的细胞质结构域相互作用,形成螺旋结构,使运输小泡附着到受体膜。小泡膜中的Rab蛋白作为小泡寻靶和融合的定时器;② 通过多个V-SNARE与靶膜T-SNRE相互作用以及ATP的水解形成预融合复合物;③ 预融合开始之后立即进行融合,但详细机理不清;④ 在融合过程中,相互作用的蛋白进行解离,如T-SNARE/V-SNARE/SNAP25相互分开,促进了进一步的融合;⑤ 含有V-SNARE小泡的形成并回到原始膜中。
